The fact that grid cells don’t map a perfect lattice is not restricted to 3D.
We can see similar differences in 2D environments after the removal of a boundary.
2 Likes
Right. But what if grid cells are in fact encoding “a place” of that “2D sheet”? In other words, you are just translating cortex-addresses to the hippocampus-addresses. You need it to replay later hc the content in the “right” place of the cortex.
I think, we had this discussion before, ![]()
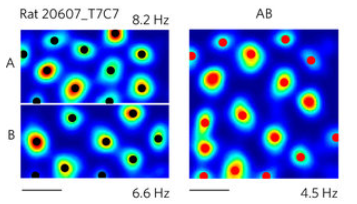
Well actually, I think that evidence supports the (Kropff & Treves, 2008) theory, which explains how grid cells form receptive fields with a “Characteristic Distance”.
Grid fields can change size and deform when a familiar environment changes size or shape.
Experience-dependent rescaling of entorhinal grids
POm, the direct sensory relay to L5a of barrel cortex, has larger RFs and poorly characterized responses compared to VPM, so it fits sending a movement direction signal. The same TC cells are also the CTC pathway though.
L6 CC cells mostly fire at the start of excitation. Maybe that’s a way to encourage activity to move around in the minicolumn. Different rates of adaptation could create a fixed order of activity moving around the minicolumn.
One study found L6 CC cells can also have long latencies before they fire. If the ones with longer latencies in response to weak input also adapt more quickly in response to strong input, they fire first when input is strong and last when input is weak. That way, activity moves around in one order when input is strong (movement in the preferred direction) and the opposite when it’s weak (movement in the opposite direction).
This wouldn’t work for long periods of time. Maybe oscillations solve that, resetting the process each cycle to form 1-d grid fields.
L6 CC cells in barrel cortex receive a very strong input from M1, maybe the strongest in all sublayers.
2 Likes
Sounds interesting, but I don’t get your link with grid cells.
Have you read this paper? L6 CC cells (excited by TC cells) gate L5 PT cells output.
https://www.cell.com/neuron/pdfExtended/S0896-6273(19)30884-0
The paper is not related to grid cells, but it could help with the role of L6 CC cells.
2 Likes
I’m not sure how the gridiness would work, now that I think about it more. My point was more that it doesn’t need to work over long periods of time because the response is gridy. It gets between equivalent points on the 1-d grid before it runs out of cells which still haven’t yet fully adapted. Cells recover from adaptation between each high-activity part of the oscillation.
No. I’ll skim it now.
During the research meeting on dec 9th 2019, @jhawkins talked about two types of sensors:
@subutai confirmed this a bit later.
Does anyone know of a neuroscience paper that discusses these two types? I tried googling for info on this, but I didn’t get far. I don’t really know how to call them or how to descibe this.
2 Likes
Start here:
You have the entire body doing inverse kinematic transform to support the gyroscope stabilization of you visual system. All the way to your supporting surface. Add to that your head and eye pointing system and you have a very good directional system.
Let me go a step further - this same system is your map back to the frame of reference “out there” and your interface between all this and your “end effectors” makes your “personal space” special as it is where your sense of agency has the greatest influence.
This whole thing is mixed in with your posture control system. If you think about it - it is really the same system.
This IS an insanely complicated system and goes directly to the point that the primary function of the brain is to run the body.
This also goes a long way towards explaining this:
Mark, this is all interesting, but where does it describe two types of sensors on our skin or retinas?
No! These senses are part of the vast somato-sensory system.
Joint angles and vestibular sensations are distinct and separate from skin and eye pattern sensations.
I could imagine someone thinking that the location of the pattern in the sensory field is the relative location but I consider that as part of the pattern.
I could be looking at this all wrong but I think that these strained efforts to put location and pattern into the single column is mostly due to a hard focus on the macro column to the exclusion of hierarchy.
I don’t think I understand. Do you mean the same physical sensor in the skin projects a signal to two different centers or regions, where one is interpreted as a fixed spacial location and another is converted into directional information?
Or is it that you disagree with the principle of having two types of sensors?
Or maybe we’re not talking about the same thing here.
This one!
1 Like
I may be off the mark, but I interpreted this comment as simply calling out two different types of nerves. Those which detect movement, and those which detect pressure (I imagine there are many more than that, such as those which detect heat, those which detect the color red, etc. etc.). The interesting thing is that Jeff mentions they travel in separate channels to the cortex, though… not sure what to make of that.
2 Likes
I found this article on mechanoreceptors, with lots of references. It goes into various types of them and how they behave. It doesn’t explicitly categorize them into the “two basic types”, but goes into the mechanics for how pressure and motion (and others) are processed.
1 Like
I am not aware of a single paper that discusses spatial vs. directionally sensitive sensors. There might be some. But there are easily found papers that discuss directionally sensitive sensors. E.g. I searched for “Motion detection in the retina”, found wikipedia article on “motion detection”, where I found this reference to Barlow and Levick 1965. I am sure there are many more recent papers out there.
Direction selective (DS) cells
“Direction selective (DS) cells in the retina are defined as neurons that respond differentially to the direction of a visual stimulus. According to Barlow and Levick (1965), the term is used to describe a group of neurons that “gives a vigorous discharge of impulses when a stimulus object is moved through its receptive field in one direction.”[23] This direction in which a set of neurons respond most strongly to is their “preferred direction”. In contrast, they do not respond at all to the opposite direction, “null direction”. The preferred direction is not dependent on the stimulus—that is, regardless of the stimulus’ size, shape, or color, the neurons respond when it is moving in their preferred direction, and do not respond if it is moving in the null direction. There are three known types of DS cells in the vertebrate retina of the mouse, ON/OFF DS ganglion cells, ON DS ganglion cells, and OFF DS ganglion cells. Each has a distinctive physiology and anatomy.”
I know that similar sensors and papers exist for somatic sensors. If I were to develop this idea further, I would do a literature search on this topic, including audition. I don’t think it would be difficult to find lots of literature.
5 Likes