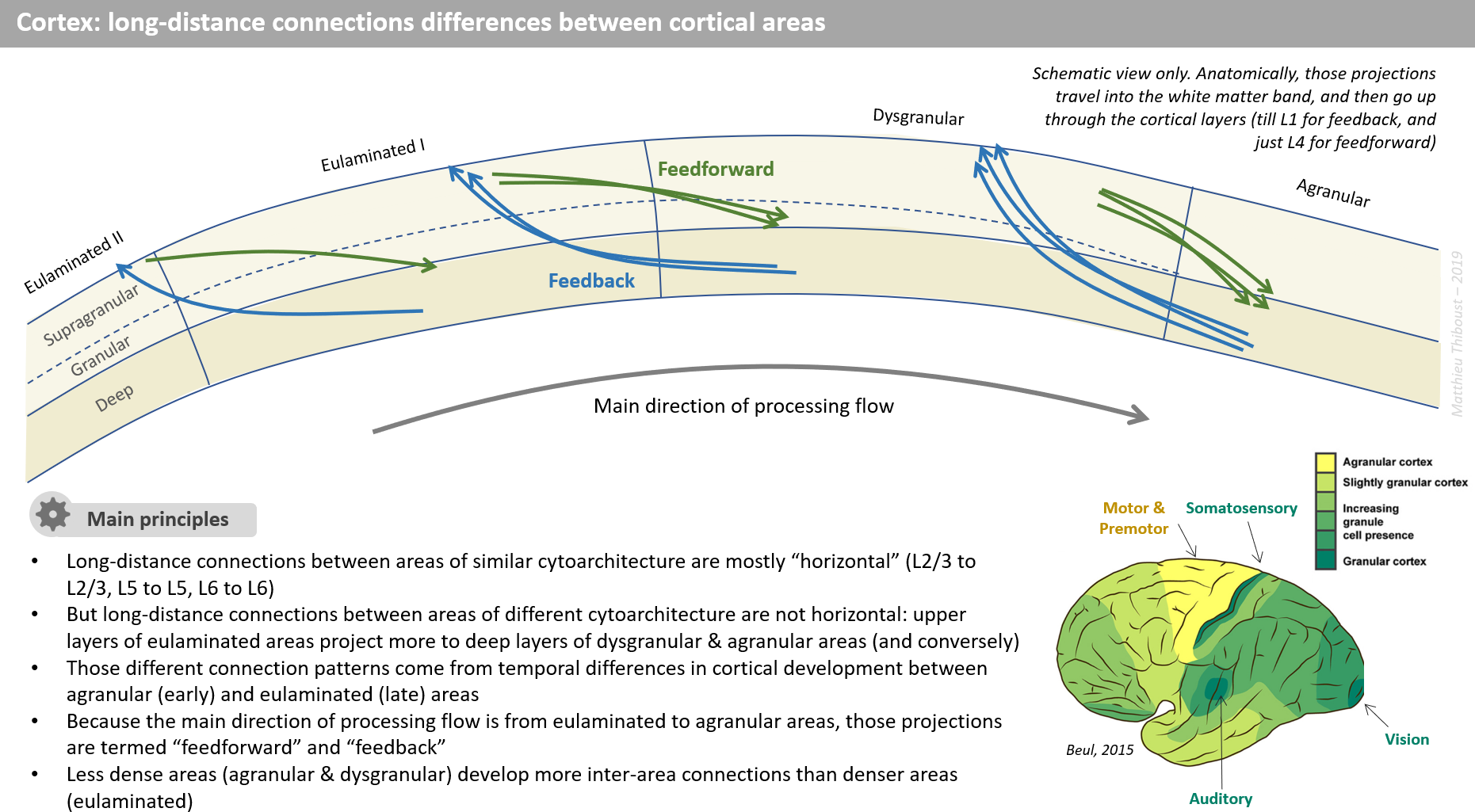
Having read a lot since the last post, I think I have now a better view of what is going on with inter and intra-area cortical connections.
I am beginning to make some order in my notes and I chose to formalize it in a visual way to make it easier to digest, and to digitalise it to share it with you. The first slides are about those cortical connections. I am planning to do the same for a bunch of other subjects I came across…
I welcome any comment on my view on the functionality behind the inter & intra cortical connections, the speculations about the nature of representation in supra vs deep layers, and the anatomical & developmental interpretation of feedfoward/feedbak.