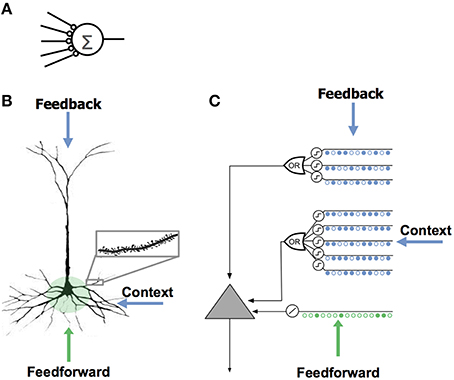
“There are three sources of input to the cell. The feedforward inputs (shown in green) which form synapses proximal to the soma, directly lead to action potentials.” Image Reference
I had the impression that a pyramidal neuron needed both feedforward and (feedback or context) excitation in order to fire an action potential. I thought this because only predicted (or depolarized) cells in a column are activated - so feedforward excitation is not enough. If feedforward excitation was enough to activate the soma then I guess all neurons in the column will activate, regardless of if they were already depolarized.
I feel I’m missing something here.
Playing with my new toy (nViz) - here is a visualization of what I mean. The left and right columns represent context and feedback neurons, while the bottom row represents feedforward inputs. The center neuron only activates when feedforward AND (context OR feedback) are provided. (segments have 70% threshold).
I believe that the lateral (context as you put it), excitation and therefore depolarization is what helps the cell to “beat out” the inhibitory cells which fire faster than the pyramidal cells, otherwise they “burst” (meaning all cells in a given column fire but they inhibit each other and so only one wins out??? – I have questions about this too). Now I’m not sure what the biological allegory for “bursting” is - or in other words, I’m not sure how the columnar cells and the inhibitory cells interact, and then are left with the “bursting” condition? But I believe at least, the answer to your question is that they are both necessary in order for there to be sparse winners (those that beat out the inhibitory cells) = but I would like some clarity on how these structures interoperate biologically, as well?
Yes, the idea is that feedforward proximal inputs are sufficient to cause a neuron to fire. But if lateral or apical input depolarizes a cell so that it fires sufficiently sooner than the others in the column, the cell will activate columnar inhibitory cells (speculated to be double bouquet cells) that inhibit the activation in the rest of the neurons in the minicolumn before those others have a chance to activate.
Thanks Scott! I have another question - does every cell in a column have proximal dendrites to the input? I initially thought that there were just one cell per column that had proximal dendrites, then the other cells in that column get excitatory input from that one ‘driver’ cell.
Good question! I can’t give a definitive answer but there’s some evidence that each cells gets FF input separately and that there are some mechanisms for ensuring the FF receptive fields for all cells in the minicolumn stay in sync. This is speculative though and you’d be better off reading the minicolumn hypothesis below and referenced papers in it.
One thing I find very interesting is that during development, it appears there are a set of cells (called subplate cells) that seem to help define the minicolumn and ensure that all the cells connect to the same FF inputs. But these subplate cells disappear afterwards so don’t appear to be able to continue performing this role later on. Very mysterious.
{kind=link}
