Sorry for disturbing.
I’m reading the paper The HTM Spatial Pooler—A Neocortical Algorithm for Online Sparse Distributed Coding by Cui.
While confused that in the part of fault Tolerance, it discuss the “receptive field center” with respectively showing the picture above. What does the “receptive field center” mean?
in the picture below, the diagram shows lines going down to an “input space.” Note the gray highlighted set of potential connections. This “receptive field” refers to the input area that the column is sampling.
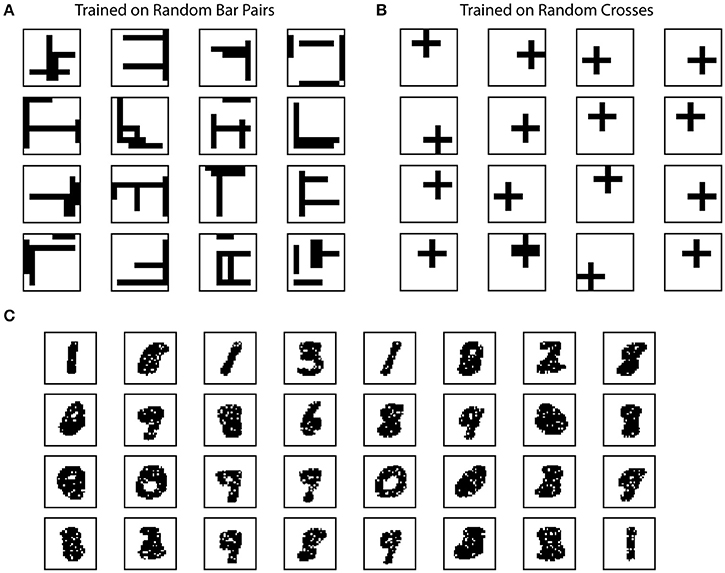
The fuller set of pictures you took this from shows that two different types of defects are being simulated. The two columns are being tested for the cases of “loss of training” or “loss of input stimulus” in this example; one is that the trained connections are eliminated (KIlled by stroke) and the other is that the inputs to the sampling space are eliminated (lesion).
Training data like this is applied before these traumatic injuries:
The accompanying text explains how the green column responds/recovers after these insults. The green column is having to use the smaller remaining set of trained inputs or retrain with new added connections depending on the type of damage.
so the location of “the receptive field center” is calculated by linear weighting the location of each neuron, which belongs to some receptive field in the Input Space, with potential connection to SP. Is it right?
One the part about the orderly arrangement of the input columns you are correct.
On the connections statement - HTM uses binary connections; either there is a connections or there is not.
I have made an “after” picture to go with the two you just posted -
the failed connections have been removed,
the two new connections shown inside the green box have been added, and
the relationship with the green box marking the input space has been added.
Thanks, Bitking! I totally understand your analysis. While just confused how the first picture (many black dots) comes, the black dots’ location means what?
(sorry, my english is so poor that hard to express my question)
The black dots are indications of sampled input points.
HTM systems tend to use less than all of the input space and still do a good job of recognizing the presented objects. That is the “sparse” part that is often referred to in HTM literature.
If a person is hiding behind a tree you don’t need to see all of him to know about where he is, and perhaps what he is wearing. These little bits that stick out around the tree are sparse information but enough; a collection of partial information can be surprisingly effective.
Can I ask two follow-up questions about this receptive field, please?
Is it correct to assume that the state of the column is the result of only a limited number of the potential connections in the receptive field (so, in the diagram, only 6 out of the 20 potential connections)?
Is it also correct that the state of the column depends on both the positive (present, black) state of a number of potential connection and the negative (absent, white) state of a number of other connections? And does this emulate how real synapses and neurons behave?
When I consider this part of HTM theory it helps me to go back to the biology that inspires the theory.
This part of the theory is modeling the apical dendrite as it snakes through the jungle of other horizontal dendrites and rising projecting axons. It will travel about 500 µm “sideways” (actually, +/- 250 µm centered on the cell body) passing though rising/projecting axons with a mean spacing of about 30 µm.
I say rising/projecting axons because some are from neighboring cells and some are projections from distant maps. To the dendrite they look like the same thing. In the picture below we can see the ascending corticocortical axon from a distant map in red. These axons do make contact at various points in the six layers but we are looking at just L1 for this discussion.
The rising axons individually contact a small local part of the dendrite, in the picture below look at the orange dot to form a mental picture of the relative scale of the cells receptive field in relation to the rising/projecting axons field of possible influence.
Please keep in mind that this picture portrays a vastly reduced number of dendrite arbors - these cells also are also spaced on 30 µm centers but if I put them all in the diagram would be to dense to see anything.
Our dendrite will not necessary have synaptic connections with all of the axons it passes - only the ones that it has learned that it has some meaningful relationship with. You know - the whole fire together/wire together thing ?
So the rectangular field portrayed in the model is actually a circular field emanating from the apex of the pyramidal cell’s soma in the biology. The rectangular shape is a simplification to make the programming easier.
The pool of potential connections corresponds to the axons closest to the path of the dendrite as it snakes through the L1 jungle. There are a much larger pool of rising axons that the dendrite is too far from to form a connection. These rising axons are within the 500 µm diameter circle centered on the cell body but no part of the cells dendrite pass close enough to form a synapse.
Other cells may have a dendrite that does make contact with this rising axon.
So - with this background lets get back to the original questions: Is it correct to assume that the state of the column is the result of only a limited number of the potential connections in the receptive field (so, in the diagram, only 6 out of the 20 potential connections)?
Yes. These correspond to the axons that the dendrite has formed a synapse with.
Is it also correct that the state of the column depends on both the positive (present, black) state of a number of potential connection and the negative (absent, white) state of a number of other connections?
Not exactly. Yes on the positive (present, black) part and no on the negative (absent, white) state. These negative location may be sampled by some other column but this particular column can’t see as these are they are just too far away from it’s dendrite.
And does this emulate how real synapses and neurons behave?
I hope that this post helps explain that HTM is a simplified version of the neuron/synapse but it does capture the essential features.
It’s baffling how much you know about this. Thank you.
Too far away? Or not connected due to negative Hebbian reïnforcement? They are within the receptive field.
The reason I asked about the negative states is because I have some confusion about some connections that cause inhibitory effects, as described in several papers. In software we can program a NOT gate/instruction, but I have trouble visualising how this may work in biology.
I heard/read @jhawkins often talk about “depolarisation”, and each time I wonder if this causes an increase in action potential or a decrease. Typically, depolarisation signifies limiting the ionisation (making a less polarised solution), and so in purely electrical terms, the action potential should decrease. But at least in a few cases, the opposite seems to be implied.
Also, I heard/saw Henry Markram once describe a type of neuron that sends out a constant output via its axon, except when its action potential is fullfilled. That would be a bio NOT gate. This is maybe a bit off-topic, but seeing the diagram you posted, for a second I thought this might be part of the mini-column structure.
Thanks - I have been studying this stuff for a long time - since the 1970s.
You are mixing different parts of neural theory in places where they don’t apply so I am not surprised that it seems confusing. I will take a stab at placing these items in the correct place and give some context in how they fit together.
In no particular order:
Position in receptive field
The reach of the individual dendrite establish the scope of the receptive field. Synapse are distributed along the dendrite and are the point where the interaction between the axon and dendrite occurs. The action of the synapses are summed along the dendrite and while the action of the synapse is binary - the action of the dendrite is to sum the total action of the synapses over a length of dendrite. Synapse closer together are more likely to initiate depolarization (see below) than those further apart. This spacing bit of neural theory is generally ignored in the HTM canon.
depolarization
This is a small but critical part of the action potential. The cell acts to pump chemicals across the cell membrane and this establishes a sort of chemical battery that makes a voltage across the cell membrane. Different inputs can lower that voltage at “hotspots” in the cell membrane. A key fact in understanding depolarization is that firing the cell is about the same as shorting out this battery at some point; this discharge spreads along the cell membrane in a traveling wave. This is the primary effect of a synapse. One synapse by itself usually can’t fire the whole nerve - it usually take a gang of them to pull this off. It is a form of voting - each one cause some depression in the voltage - depolarization. It there is enough this starts the wave called the action potential.The closer together or the closer to the soma, the more effect a synapse can exert. Quoting George Orwell: “All animals are equal, but some animals are more equal than others.” This works for synapses too!.
*Action Potential
I really can’t do better than this wiki entry:
Neurotransmitters.
One of the oldest preserved functions from our origins as layers of pond scum is our use of chemical messengers. This predates defined neural structures and for that matter was well before cell walls were a thing. These messengers have been improved and incorporated into the nervous system as it has evolved.
A rouges gallery include many different chemicals and methods of action.
Neurotransmitters that fall into the category of amino acids
✦ Glutamate
✦ Aspartate
✦ Glycine
✦ D-serine
✦ Gamma-aminobutyric acid (GABA)
Monoamines or other biogenic amines considered as neurotransmitters
✦ Serotonin
✦ Norepinephrine
✦ Epinephrine
✦ Histamine
✦ Melatonin
Neurotransmitters that fall into the category of peptides
✦ Beta-endorphin
✦ Opioid peptides
✦ Somatostatin
✦ Calcitonin
✦ Vasopressin
✦ Oxytocin
✦ Glucagon
» Apart from these, there are several other important neurotransmitters, such as acetylcholine, dopamine, adenosine, and nitric oxide. So far, about 50 neuroactive peptides have been discovered.
» Sometimes, neurotransmitters are also classified as excitatory and inhibitory. This classification is based on their actions on the neurons. Excitatory neurotransmitters are those that excite the neurons and stimulate the brain, while inhibitory neurotransmitters are known for having a calming effect on the brain.
» Neurotransmitters like, GABA and serotonin come under the category of inhibitory neurotransmitters, while epinephrine and norepinephrine are the excitatory neurotransmitters. Dopamine on the other hand, can act as an excitatory, as well as an inhibitory neurotransmitter.
» However, the effect of a neurotransmitter on a postsynaptic cell depends on the receptors present in it. For some neurotransmitters, like glutamate, the important receptors have excitatory properties. On the other hand, most of the important receptors produce an inhibitory effect for GABA. But there are some neurotransmitters, for which both types of receptors exist.
It is very hard to make sweeping statement on how these work.
Some of these are passed from cells at the cleft between synapse and axon.
Some are circulated in the bloodstream.
Some are released into the inter-cellular space.
Some are fast acting, some sort of “set the mood.”
Some leave a longer term effect by either enhancing or suppressing future firing of the target neuron. The time scale varies depending on the exact mechanism of action.
This wiki page is generally correct but is very specific on what is considers a neural chemical messenger. The brain is not so neat about these things.
Inhibitory vs excitatory
Let’s try to separate what is going on in “Hebbian learning” and “chemical signalling.”
As I stated - the action of the chemical messenger can either facilitate depolarization or inhibit it. This effect can range from a mild influence to initiation of immediate effect.
The cells used in in the HTM theory are of both excitatory and inhibitory types, but the are not mixed together in the same parts of the system. This thread is focused on the SP part of the theory but in the larger HTM universe we use both types of cells.
The cells used in SP are excitatory when summing inputs. As the columns are summing the input field these triggers the inter-neurons to form an inhibitory effect where the strongest responding column suppresses the action of more weakly resounding columns. HTM uses the MAX function (k-winner) to select the strongest column in an area and suppresses the rest; in the biology this is performed with interactions with inhibitory inter-neurons. In both cases this enforces sparsity between macro-columns. The resulting effects are similar but not exactly the same.
In the TP portion of the theory, the mini-column that has been primed by a partial depolarization in the previous cycle fires faster and triggers shorter range inhibitory inter-neurons to suppress the rest of the macro-columm. Ya - prediction!
Hebbian learning
Hebbian learning is a proposed mechanism that relates the firing timing between two cells and the effect on the long term response of the receptive cell. Learning is a different thing from a nerve just firing. A cell firing is a one time event that transforms whatever influences are acting (Inhibitory or excitatory) on a cell into an action potential event. Learning is a long term change in the cell - normally thought to be some change in synapses - that changes the cells response to the influences to the cell. This influence could act to grow or shrink the strength of a synaptic connection.
Positive/Negative Hebbian reinforcement.
The simplest description is that if the cells fire at about the same time in a certain order the synapse becomes stronger and if the order is reversed the synapse becomes less effective. If this synapse is not used over a long time this synapse loses effectiveness (gets weaker/forgets) @Falco - The effect of the spatial pooler input is always excitatory but it is more or less so based on this learning. There is no negative input or summation.
This has very little to do with the type of the neurotransmitter involved and it’s nominal Inhibitory vs excitatory effects. The outcome if this factoid is that the some of the input bits may be the compliment of the expected polarity of the input channel. That is outside scope of the Spatial Pooler and more correctly associated with the nature of the signal being sampled. (the encoder)
@Bitking, I appreciate what you’re trying to do, but this is too much. Writing a 3-page essay to answer a question is not productive. 95% of what you wrote I already knew from reading On Intelligence, watching HTM school and the Khan Academy video’s. You lost half an hour of your life writing up this stuff, and honnestly, it made me feel like an idiot. Seriously, I’m almost afraid posting questions now.
That’s what I understood too, but then we’re dealing with a simple misnomer. This effect should be called polarisation. The effect increases the difference in electrical charge by increasing the concentration of ions in the plasma solution at one side of the cell membrane. (The positive and negative poles across the membrane increase). Only when enough synapses act in concert, is the charge high enough so that a discharge can happen. Which I guess causes a partial depolarisation as the soma fires.
So, it’s polarisation by the individual synapses, and depolarisation by the soma after enough synapses have transmitted their input.
As for the inhibitory effect, I’m almost certain I read in more than one place in Numenta documents that distal and/or apical dendrites fire to keep neighboring cells from becoming active. (These can’t have been purely chemical effects, since that would have been too slow). I can’t look it up right now, but I wouldn’t be surprised if that is the mechanism that makes sure only one mini-column becomes active, even though several mini-columns have connections with positive potential connections from the receptive field (black dots in the diagrams). I still have to read the two newest papers. Maybe I’ll find more information there.
Sorry about that. I write in part to refresh my memory on these topics; it is necessary to retain the basics. Everyone should consider mentoring for this reason. These BAMI questions are very far from where I am working and I need the break sometimes.
I came to HTM after years of reading biology and neural network lore. There are numerous things about HTM that I have had to fill in or frankly - just ignore - because they hammer the biology to fit the theory.
On the (de)polarization thing: there are all kind of things in neurology that are not named in ways that make any sense to me. And don’t get me started on Ben Franklin and polarity.
@Falco - “As for the inhibitory effect, I’m almost certain I read in more than one place in Numenta documents that distal and/or apical dendrites fire to keep neighboring cells from becoming active.”
They don’t. Not even a little. It is the inhibitory inter-neurons that do this.
I think you need to read outside of the HTM literature.
HTM is not very keen on anything other than the basic pyramidal cells. The inhibitory inter-neurons get a casual mention but they are definitely second class citizens in the zoo of cells in the cortex - at least in the HTM canon. I do try to restrict my answer here to the basic way that HTM uses the biology to make things a bit easier to follow but only to a certain point. At some point the HTM theory will have to be extended to include the rest of the cells in the cortex.
Yesterday night, after committing my post, I felt bad. I thought I had been insulting. I thought you might get angry at me, and this thread would escalate. I even feared I might having to stop posting on this forum.
And here I see, the first thing you wrote is “Sorry”…
I’m the one who should apologize. I was tired and a bit irritable (which is no excuse), and I should have understood that you made a great effort in helping me understand this complex material. How were you to know what I have read and what not. What is understood and what not.
I’m sorry @Bitking. I was out of line. It’s refreshing to find a forum where people think before they write, and where posts are fueled by knowledge instead of ego.
Ok. Would you perhaps know of a thread on the forum where this is discussed? We’re getting a bit too far off-topic, and I think this is important. (And definitely something I don’t know). Is it possible that the effect of these inter-neurons can be simplified within HTM without programming specific gates or objects to include their behavior within the theory?
Well, it is a work-in-progress after all. As a teacher of mine used to quib: Rome and Londen didn’t burn down in one night. ;-).
Your question is very “on topic” for the spatial pooler paper and the fact that you have questions suggests that others may have the same questions. The “sparsing via the inhibition” is not explained very well in the original paper and in the 19 times that the word is mentioned the text does not go very far into explaining how it works in the biology - only that somehow it does. On the plus side - the reference section does point to some good papers.
In the SP paper the inhibition is mentioned in figure 1: "A local inhibition mechanism ensures that a small fraction of the SP mini-columns that receive most of the inputs are active within the local inhibition radius (shaded blue circle).
This is the hand-off from local inhibition to the k-winner-take-all approximation: " The SP models local inhibition among neighboring mini-columns. This inhibition implements a k -winners-take-all computation (Majani et al., 1988; Makhzani and Frey, 2015). At any time, only a small fraction of the mini-columns with the most active inputs become active."
From the forum:
From the linked SP paper:
In the SP paper reference section linked papers discuss the inhibitory inter-neurons in depth.
Porter, J. T., Johnson, C. K., and Agmon, A. (2001). Diverse types of interneurons generate thalamus-evoked feedforward inhibition in the mouse barrel cortex. J. Neurosci. 21, 2699–2710.
Markram, H., Toledo-Rodriguez, M., Wang, Y., Gupta, A., Silberberg, G., and Wu, C. (2004). Interneurons of the neocortical inhibitory system. Nat. Rev. Neurosci. 5, 793–807. doi: 10.1038/nrn1519
Swadlow, H. A. (2002). Thalamocortical control of feed-forward inhibition in awake somatosensory “barrel” cortex. Philos. Trans. R. Soc. Lond. B Biol. Sci. 357, 1717–1727. doi: 10.1098/rstb.2002.1156
The inter-neuron function is approximated by the k-winner-take-all function:
Majani, E., Erlanson, R., and Abu-Mostafa, Y. (1988). “On the K-winners-take-all network,” in Proceedings of the First International Conference on Neural Information Processing Systems (Lake Tahoe, NV), 634–642.
@Falco@Bitking I just read through your most recent interactions and I must commend you both for being shining examples of civil discussion. This is a very high-quality thread IMO for new readers.